
臨海管道微生物腐蝕損傷機制與防護
2021-10-29 17:28:29
作者:李光泉,李廣芳,,王俊強,張天遂,,張斐,蔣習民,劉宏芳 來源:中國腐蝕與防護學報
分享至:


摘要
海洋油氣管道作為大量、長距離輸送油氣資源最主要的方式,擔負著海上油氣集輸的重要任務,也被稱為海洋油氣工程的“生命線”。然而海洋環境下的微生物腐蝕是造成海洋油氣管道腐蝕損傷的重要原因之一。本文結合海洋油氣輸送管道的服役環境,綜述了海洋環境下臨海管道微生物腐蝕失效的研究進展,重點介紹了有代表性的硫酸鹽還原菌和鐵氧化菌在海洋環境下引起的微生物腐蝕規律和機理,并在此基礎上總結了相應的海洋管道防護方法,為微生物腐蝕損傷的研究及控制提供參考。
關鍵詞: 臨海管道 ; 微生物腐蝕 ; 腐蝕防護
海洋中的管線,比起陸地環境中的而言,腐蝕更為嚴重,而且不易維修。海洋管道腐蝕不僅僅造成材料的浪費、環境的污染,更嚴重地是因腐蝕造成的油、氣泄露引發災難性事故。據統計,在海底管道發生失效的事故中,腐蝕引起的事故約占50%的比例,每年因腐蝕帶來的經濟損失約占各個國家國民生產總值的2%~4%。因此,在海洋石油開發領域,海洋腐蝕問題給人類帶來了巨大的困擾,海上石油開發過程中的管道腐蝕與防護問題亟待解決[1]。
由于海洋環境復雜,對于海洋管道鋼鐵材料而言,除了受到海水的化學腐蝕、電化學腐蝕,還受到海洋微生物、生物的附著和污損[2-4]。海水環境中微生物腐蝕 (MIC) 最為普遍,與其相關的腐蝕失效和破壞已達到涉海材料總量的20%[5]。目前海洋環境已發現眾多微生物菌群,包括古菌、病毒、酵母菌、放線菌、真菌以及與腐蝕相關的各種細菌[6],約占全球微生物數量的13%。研究表明微生物的菌落結構及活性均會隨著海水的深度的變化而變化,如臨海區、淺海區等氧氣含量豐富的位置,藻類[7]、海洋生物、鐵氧化細菌、霉菌等引起的海洋污損嚴重,而在海泥區,以硫酸鹽還原菌 (SRB) 為主的厭氧微生物會引發腐蝕過程的發生。在我國遼東灣以及南海珠江口東部海底沉積物中SRB含量高達460、913個/克濕泥。在海洋底泥中,尤其是在低溶解氧條件下,高含量的SRB明顯加速金屬材料的腐蝕速度。
我國的海洋輸送管線材料主要包括兩類:金屬材料和非金屬材料,其中海洋油氣管線主要以金屬管道為主。根據實際應用條件的不同,金屬管線材料分為碳鋼管道、不銹鋼管和銅鎳合金管等。碳鋼管線材料由于其成本低廉,力學性能優良從而在我國被廣泛應用,碳鋼管道中X65、X70、X80管線鋼是油氣管道常用的材料。此外,X100和X120等新型超高強度的管線鋼也在工程中逐步應用[8]。不銹鋼雖然具有良好的耐蝕性能,但海水中的點蝕穿孔現象以及高昂的成本往往限制了部分不銹鋼的應用。銅及銅鎳合金管道被普遍認為是兼具了耐海水腐蝕和耐海洋生物腐蝕的優良性能。然而一些研究卻表明銅及其合金材料并不能有效防止SRB引起的腐蝕[9],銅在SRB存在的條件下反而會遭受嚴重的微生物腐蝕。因此,對于實際應用中的海洋油氣輸送管線受到的海洋微生物腐蝕尤其是SRB造成的腐蝕研究非常重要。
因此,為保證管道的安全運作,研究海水環境中管道的腐蝕特征、實時檢測管道腐蝕狀態,己成為亟待解決的焦點問題[10,11]。鑒于海洋管線腐蝕的危害性與海洋環境的復雜性,研究海洋鋼鐵材料的微生物腐蝕機制具有重要意義。近年來,人們從微觀和宏觀兩個角度,對微生物腐蝕特征、機制等做了大量的研究。此工作旨在系統總結臨海管道腐蝕特征與機制,重點討論臨海環境微生物腐蝕機制,總結目前海洋腐蝕與防護研究所面臨的挑戰,并對未來的發展方向進行展望,為更好地認識微生物污損行為和開發環保型方法提供理論支撐。
1 臨海環境海洋腐蝕特征及微生物腐蝕規律
一般說來,臨海環境海洋腐蝕主要集中在以下幾方面:(1) 海洋大氣環境腐蝕。該腐蝕環境為高鹽含量、高濕度的大氣環境,且氧氣供應充分,但該區域微生物腐蝕的影響較少。(2) 海水飛濺干濕交替的環境腐蝕。在高潮線以上的海水飛濺區受波浪與潮汐作用,海水飛濺攜帶的腐蝕性離子在干熱交替環境下易對鋼結構造成較強的腐蝕作用。此區域供氧充分,導致腐蝕加劇,腐蝕率遠遠高于全浸區或海洋大氣區。(3) 海洋潮差區腐蝕。海洋潮差區的腐蝕與海水的飛濺區不同,雖然潮差區也屬于干濕交替的腐蝕環境,但由于潮水的漲落使得潮差區的干濕交替具有穩定的周期。這種周期性的干濕交替也會使得海水中某些好氧微生物周期性的附著在金屬表面。(4) 全浸區腐蝕。海洋全浸區是指海水最低潮位下0.2~2 m的區域,也是微生物腐蝕最活躍的區域之一。材料在此區域不僅受到海水溶解氧、鹽度、溫度、流速、pH等各類環境因素的腐蝕影響,海水中大量的好氧菌如鐵氧化細菌、銅綠假單胞菌等也會對該區域金屬設備造成極大腐蝕破壞,微生物污損作用不容忽視[12]。(5) 海泥區腐蝕。海泥區一般被認為是海洋環境中腐蝕情況較輕微的區域,然而由于其低含氧量和富營養成分,海泥區為厭氧菌如硫酸鹽還原菌等提供了良好的生存環境。在底泥表層 (底泥和海水界面) 處由于厭氧微生物作用導致的微生物腐蝕會加速附近位置基體管線材料腐蝕失效。特別是海陸交匯環境,海水和淡水交界位置,是厭氧微生物硫酸鹽還原菌生長繁殖旺盛的位置,微生物腐蝕是關鍵失效原因之一[13-15]。
2 海洋管線腐蝕類型及腐蝕機制
海底管道輸送的介質主要有原油、天然氣、地層水及其混合物,海底管線主要腐蝕類型大體分為內腐蝕和外腐蝕兩種。內腐蝕的主要包括CO2/H2S酸性氣體腐蝕、垢下腐蝕、微生物腐蝕等;外腐蝕主要來源于海底管道所處的海水環境,主要有海水腐蝕和海底底泥腐蝕。海底管線內外受到各種形式的腐蝕破壞,本文將概述常見典型的微生物腐蝕、縫隙腐蝕、應力腐蝕開裂,及多種腐蝕現象并存并相互促進的腐蝕失效模式。
2.1 微生物腐蝕
海洋環境中與腐蝕相關的微生物主要包括細菌、真菌及藻類,尤其以細菌為主。腐蝕性細菌又分為好氧菌和厭氧菌,前者主要包括鐵代謝菌 (IOB),后者主要指SRB[16]。海洋環境中SRB、IOB是普遍存在的,研究其造成的金屬腐蝕具有典型性,可以得到許多具有普適性的觀點和結論。而目前的研究表明,SRB在缺氧環境中,是最具有腐蝕性的關鍵微生物種類之一[17]。IOB好氧細菌主要活躍在富氧的海水區,除了其自身對基體材料造成的腐蝕,通常和SRB具有協同作用,加速金屬的腐蝕。
2.1.1 硫酸鹽還原菌的腐蝕機理
常壓下在海水區與底泥區有關SRB對金屬的腐蝕機理主要包括以下3種:
(1) 陰極去極化理論。該理論認為金屬作為陽極失去電子被腐蝕,介質中的質子得到電子在金屬表面產生H,SRB在氫化酶作用下消耗H,從而在腐蝕過程中促進陰極去極化的作用,加速腐蝕。涉及如下反應:
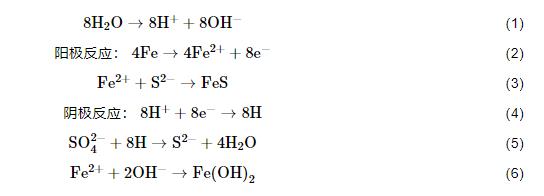
總反應:
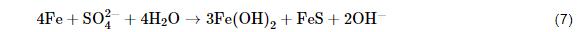
(2) 代謝產物去極化理論。傳統的腐蝕理論認為微生物并未直接參與到金屬腐蝕過程中,而是微生物的代謝產物對金屬產生腐蝕作用。SRB腐蝕的發生與生物膜密不可分,生物膜在金屬材料表面吸附和生長在非常短時間內就可以完成,生物膜主要有微生物細胞體、代謝產生的胞外聚合物、腐蝕產物以及少量吸附的有機和無機物組成。Craig等[18]認為,細菌代謝產物的去極化包括H2S和FeS去極化。厭氧環境下,陰極產生H2S,從而使腐蝕加快,陽極反應也因FeS的生成而加快金屬腐蝕。除了無機代謝物對腐蝕的影響,代謝產生的胞外聚合物在SRB腐蝕過程中也發揮重要作用。胞外聚合物是生物膜的主要組成成分,包括莢膜、粘液層、及其他表面物質,其有機部分主要由多聚糖、蛋白質和少量DNA、脂和腐植酸組成。劉宏芳研究團隊[19]發現常壓下培養14 d SRB產生的胞外聚合物量最多,而且低濃度的胞外聚合物 (EPS) 對碳鋼具有抑制作用,而高濃度的對碳鋼的腐蝕具有促進作用。Stadler等[20]發現SRB代謝產生的胞外聚合物可以明顯的抑制碳鋼的腐蝕,而且胞外聚合物的附著促進了更多SRB吸附于碳鋼表面。Ghafari等[21]從底泥和溫泉水中分離了21種細菌,研究發現僅有兩種細菌產生的胞外聚合物對碳鋼具有良好的緩蝕作用。由此可見,胞外聚合物與碳鋼腐蝕之間的內在關系與微生物的濃度以及種類有著密切關系。然而,EPS引起的腐蝕機制仍然需要進一步系統研究。雖然有報道提出EPS中的某些組分具有較高的還原電位,具備一定的氧化能力,其自身的還原可以與陽極鐵氧化構成電極對,從而加速試樣的腐蝕過程,然而具體的活性物種尚不清楚[22]。Jin等[23]發現,適量的Fe濃度可以促進生物膜和EPS形成,而且EPS中部分官能團可以與鐵離子結合,促進陽極溶解,進而加速腐蝕[22],該發現一定程度上為揭示EPS促進Fe的陽極溶解過程提供了理論支撐。
(3) 生物陰極催化 (生物電化學機理)。以上討論的理論雖然在一定程度上解釋了SRB的微生物腐蝕過程,但是并未明確SRB在腐蝕中所起的作用。隨著研究的不斷深入,人們慢慢發現了這些理論的局限性[24]。例如,Xu等[25]提出傳統的陰極去極化理論并不能適用于所有SRB的腐蝕現象,有很多并不能分泌氫化酶的SRB仍然會對金屬產生腐蝕。從微生物能量學的角度出發,維持SRB生命活動不僅消耗ATP,還需要電子[26]。他們基于“饑餓”實驗,相比較于碳源 (電子供體) 充足的環境,在沒有碳源或者很少量碳源存在的條件下,發生了更嚴重的微生物腐蝕[27]。基于此,他們認為SRB生物膜通過從基體金屬材料中獲取能量來獲得維持其生命活動,提出了生物陰極催化硫酸鹽還原機理 (BCSR)[28,29],如圖1所示。BCSR理論認為在周圍環境中有機碳源充足時,SRB首先利用有機物質作為電子供體。但是貼近金屬表面的底層SRB生物被膜由于受到擴散和上層生物被膜消耗碳源的限制,導致其無法有效從外界獲取碳源。為了獲得維持其生存的能量,金屬成為唯一電子供體[28],底層生物被膜從金屬中捕獲電子來進行厭氧呼吸、還原硫酸根,獲取氧化還原過程中釋放的能量,從而導致了嚴重的金屬腐蝕[29]。文獻[30,31]同樣觀察到長期處于無碳源環境下的SRB造成了更為嚴重的點蝕現象。生物陰極催化機制為微生物腐蝕研究提供了新的切入點,目前被認為是SRB促進金屬局部腐蝕的主要原因之一。此外,BCSR理論的提出不僅清楚的闡釋了硫酸鹽還原菌腐蝕的具體過程,還解決了長期以來一直存在于微生物腐蝕領域中的一個問題:微生物為什么要腐蝕金屬。BCSR理論還可以解釋海洋環境下其它細菌對金屬的腐蝕過程,如硝酸鹽還原菌和古細菌同樣是為了獲得維持生存的能量而腐蝕金屬[32]。
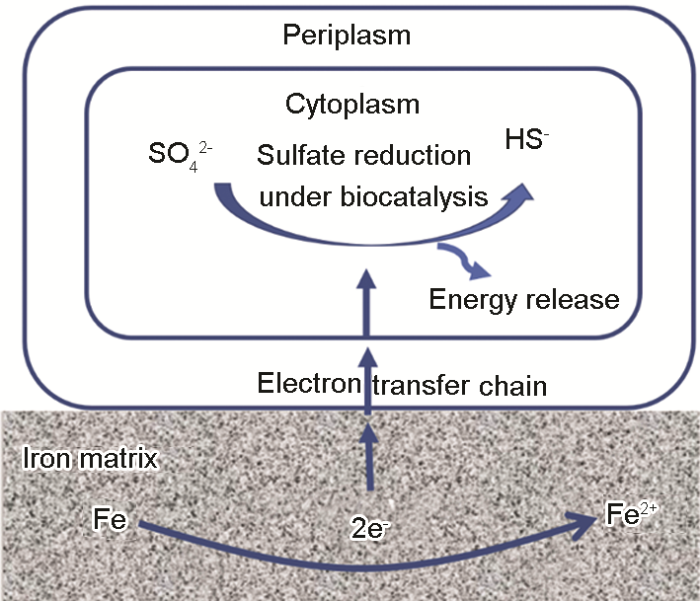
圖1 生物陰極催化硫酸鹽還原機理圖[28,29]
進一步了解BCSR作用機理,生物膜的導電性以及細胞外的電子傳遞問題成為廣泛關注的問題。Torres等[33]提出細胞外電子傳遞(EET) 的主要途徑有直接電子傳遞與間接電子傳遞兩種方式。直接電子傳遞主要依靠細菌細胞膜上的導電蛋白以及細菌自身與鞭毛類似的導電納米線 (Pili)[34];而間接電子傳遞是通過細菌自身分泌的可溶性氧化還原分子進行電子傳遞。Xu和Gu[27]首次將胞外電子傳遞的概念引入到微生物腐蝕研究中,從生物電化學和生物能量學角度來解釋微生物為什么以及如何造成金屬腐蝕。圖2描述了3種細胞外電子傳遞的方式,這3種傳遞方式并不是單一存在的,至于哪種傳遞方式發揮主導作用,要視具體微生物環境來定[16]。基因工程方法可抑制某些特定基因的蛋白和酶的表達,從而研究MIC中的電子傳遞載體,尋找抑制微生物腐蝕的靶點和方法,指導抗菌劑的研發、輸油管道的材料選擇以及抗菌材料的發展。然而海洋環境壓力和溫度的隨機變化也有可能發生改變SRB代謝機制和對金屬材料的腐蝕機制,目前尚缺乏相應的研究工作。
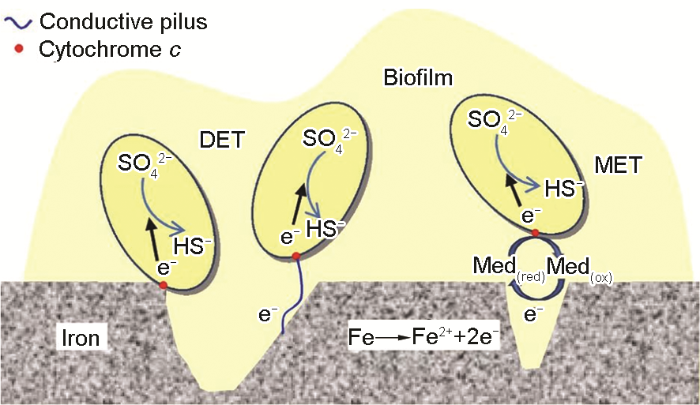
圖2 SRB腐蝕金屬過程中的電子轉移方式[16]
2.1.2 好氧細菌腐蝕機制
IOB是一類以O2作為最終電子受體,通過氧化Fe2+至Fe3+獲得能量生長的微生物,屬于典型的好氧腐蝕菌[35]。除IOB外,銅綠假單胞菌也是一種能夠引起碳鋼腐蝕的常見好氧菌[12]。目前較普遍的觀點認為典型的碳鋼點蝕機理是由IOB腐蝕產物鐵氧化合物引起的縫隙腐蝕[36]。例如,劉宏偉和劉宏芳[37]研究了IOB對Q235鋼的腐蝕狀況,結果表明,在空白無IOB存在條件下,試樣腐蝕較輕且主要表現為均勻腐蝕,而在IOB存在的條件下,試樣腐蝕加速且產生明顯的點蝕坑。IOB對金屬材料的整個腐蝕電化學過程中的化學反應式如下所示[38]:
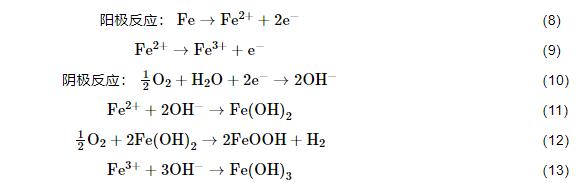
圖3描述了由鐵氧化合物引起的縫隙腐蝕機理圖[36],好氧條件下,鐵氧化合物膜下碳鋼基底會形成很多小的陽極活性位點,陽極鐵失去電子,陰極氧氣被還原,氧的去極化過程會生產OH-,進而會產生鐵氧化合物 (如FeOOH、Fe(OH)3等),鐵氧化合物的形成又進一步促進陽極的溶解,從而會加速點蝕的形成。
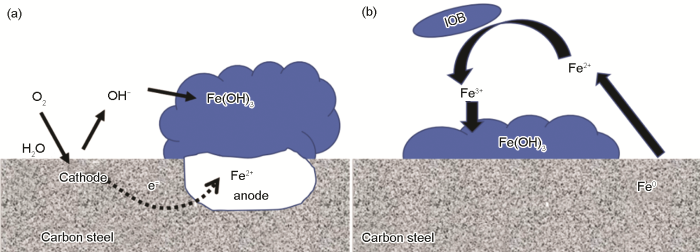
圖3 由Fe(OH)3沉淀形成的點蝕機理和IOB縫隙腐蝕機理示意圖[36]
2.1.3 混合細菌協同腐蝕
實際海洋環境復雜多變,種類繁多的微生物在海水中共同存在,它們彼此拮抗或促進,相比于單獨存在的情況,微生物的代謝增殖特點必然會發生改變。在溶解氧豐富的海水區,好氧與厭氧菌之間的協同作用是造成金屬腐蝕的一大主要關鍵因素。好氧微生物可改變環境參數,為厭氧菌的存活代謝提供局部厭氧環境。厭氧的SRB和好氧的IOB是目前公認的兩大腐蝕性微生物,二者共同存在條件下的腐蝕行為和機理研究具有典型代表性,與實際情況更為相符。劉宏芳課題組[39]系統研究了在IOB與SRB共存條件下的碳鋼腐蝕失效行為。研究認為,與單純的SRB存在條件下相比較,混合菌共存產生更多含硫的腐蝕產物,說明IOB的存在促進SRB的生長與繁衍,導致更為嚴重的點蝕發生,腐蝕機理如圖4所示。IOB與SRB首先附著在材料基底表面 (圖4a);在有限的溶氧量條件下,IOB優先通過代謝增殖形成生物膜以及鐵的氧化物沉積在材料基體表面,創建一個適合SRB存活的相對封閉的微環境,IOB消耗氧氣的代謝活動為SRB的代謝繁殖所需的厭氧環境進一步優化了條件 (圖4b)。除此之外,IOB的代謝產物鐵氧化合物的沉積進一步鞏固SRB生長所需的厭氧微環境并促進金屬腐蝕的發生。生物膜的存在阻隔了SRB與外界碳源的接觸,SRB直接從金屬中獲取電子維持生理活動所需的能量,最終導致點蝕的發生 (圖4c)。總體來說,IOB與SRB的協同腐蝕的過程中,好氧菌首先為厭氧菌創造厭氧微環境并產生點蝕,繼而厭氧菌進一步誘導并加劇腐蝕。這種協同作用可以能夠合理地解釋油田管線中混合細菌引起的局部穿孔現象。
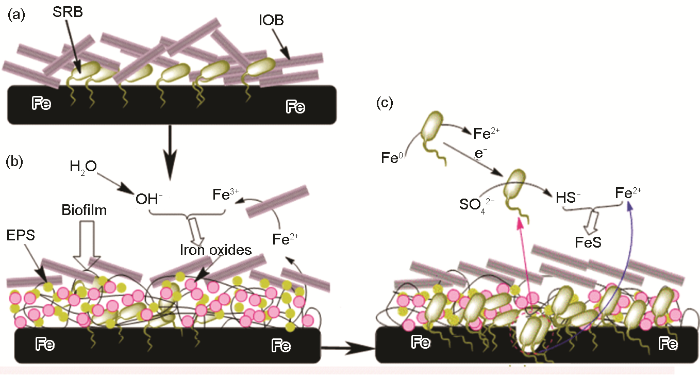
圖4 SRB-IOB混合菌誘導金屬點蝕機理示意圖
2.2 應力腐蝕開裂
2.2.1 應力腐蝕開裂機理
臨海管線面臨著第二個典型的腐蝕問題為應力腐蝕 (SCC)。應力腐蝕開裂是一種比較嚴重的腐蝕形式,初級裂紋是在環境與應力協同作用下孕育形成,接著裂紋擴展并不斷萌生次級裂紋,最終在沒有明顯的預兆下突然發生脆性斷裂。海底油氣集輸管線在運行過程中必然承受著巨大的應力,從而造成應力腐蝕,應力腐蝕開裂是造成裝備失效的主要原因之一,是油氣輸送管線運行過程中破壞性和危害性最大的一種腐蝕。應力腐蝕機理包括經典的陽極溶解機理與氫致開裂機理。傳統的陽極溶解SCC機理,認為金屬材料在應力和腐蝕的協同作用下,局部位置產生微裂紋,而裂紋尖端的快速陽極溶解引起應力腐蝕開裂[40]。氫致開裂理論則認為腐蝕開裂是在氫和應力的共同作用而導致金屬材料產生脆性斷裂,即氫脆。和常壓環境相比,海洋環境中由于溶解氧濃度的減少,陰極反應會逐漸由吸氧反應向析氫反應轉變,在金屬表面發生陰極析氫反應,造成H在鋼鐵表面的吸附及向內部的擴散,由于氫原子很小,很容易在金屬的晶格中移動,使鋼鐵結構脆變,同時在交變載荷作用下,鋼鐵結構很容易發生脆性斷裂。同時,微生物誘導腐蝕也能產生氫源,促進金屬氫致開裂的發生。另外一方面析氫反應產生的氫氣由于氣壓的存在會進一步促進涂層的剝離[41]。
2.2.2 SRB誘導應力腐蝕開裂
現場失效分析表明,SRB的活動對基體材料應力腐蝕敏感性及應力腐蝕開裂均有一定的影響。在海底泥土中,SRB的存在對應力腐蝕開裂的影響表現為點蝕的發生。由于SRB會在碳鋼表面形成疏松多孔的生物膜,生物膜中除了包含大量有毒的細菌代謝產物,還夾帶H+、Cl-、H2S,均會在碳鋼表面誘發并產生大量點蝕[42]。點蝕坑外加應力作用便能促進次生裂紋的產生及裂紋擴展,最終發生腐蝕開裂[43]。研究發現H2S分壓增加會加速H原子在金屬中的擴散,提高了金屬發生氫致開裂的敏感性[44],另一方面,SRB存在下,使得局部環境呈現較低pH值,應力腐蝕開裂受陽極溶解和氫致開裂機理混合控制。文獻[45]研究認為X80管線鋼在應力腐蝕行為與SRB活性密切相關,一定含量的SRB可加速陽極溶解、硫化物的形成,與氫致開裂共同作用導致混合斷裂。Biezma[46]研究認為SCC裂紋萌生和擴展速率取決于SRB的數量。Wu等[47]通過“有無加載應力和有無接種SRB”交互作用的實驗觀察到,加載應力與SRB的活動均可以改變生物膜特性,增大腐蝕敏感性,SRB誘導的點蝕是管線鋼應力腐蝕開裂的直接原因。盡管大量研究已表明SRB對土壤環境、海洋環境中的鋼結構材料的應力腐蝕表現出促進作用,然而SRB協同應力腐蝕的機理還尚且沒有一個統一的定論,仍需要進一步研究。
2.3 縫隙腐蝕
臨海管線面臨著第三個關鍵腐蝕問題是臨海管道裸漏后或補口損傷后,剝離涂層導致的縫隙腐蝕。沉積物覆蓋導致的縫隙腐蝕是工業界非常常見的一種局部腐蝕,也是造成管線穿孔的主要原因之一。縫隙腐蝕包括孕育、開始和發展三個階段,這三個階段的發展速度又取決于縫隙的幾何尺寸 (縫隙寬度、縫隙深度及縫隙內外面積比) 和環境條件 (溶解氧濃度、pH、Cl-濃度等)。縫隙腐蝕的機制主要包括氧濃差電池機制和IR降理論,這兩種理論分別源于縫隙內外水化學特性的差異和縫隙內外的電位差。發生縫隙腐蝕的縫隙寬度在0.025~0.1 mm,該寬度足以使深海環境中高濃度Cl-進入并滯留在縫隙中,參與到縫隙內的腐蝕過程,促進局部腐蝕。氧濃差電池機制認為,在腐蝕孕育階段,隨著縫隙內金屬的溶解,縫隙內O2被不斷消耗,縫隙內正電荷增加,為了保持電中性,Cl-等進入縫隙內發生反應使溶液酸化,導致縫隙內溶液pH值降低而進一步加劇縫隙內金屬溶解,上述過程反復進行最終造成嚴重的縫隙腐蝕。因此,氧濃差電池是此種機制下的腐蝕起因,而閉塞電池的酸化自催化作用則是造成縫隙腐蝕加速進行的根本原因。氧濃差電池機制是氧濃差與自催化效應共同作用下造成的縫隙腐蝕。IR降理論認為縫隙內O2被消耗殆盡后,縫內陽極區和縫隙外陰極區之間會出現很長的電流通路,若此通路流經的溶液介質電阻為R,當電流I流經此介質時會產生一個IR電壓降,這個電壓降導致縫隙內和縫隙外產生電位差,從而導致縫隙腐蝕發生[48,49]。此外,大小在1~2 μm的SRB同樣可以進入縫隙,參與金屬腐蝕。然而,目前SRB與縫隙腐蝕協同機制尚不清楚,需要開展針對性研究。涂層的剝離是一個緩慢生長的過程,深海環境中Cl-濃度較高,因此SRB、縫隙的幾何尺寸和縫隙內水化學特性是影響剝離涂層下金屬腐蝕的主要因素。在低溫高壓環境中,這些影響因素對于金屬腐蝕發揮的關鍵作用需要進一步探明,對于剝離涂層下的金屬管線與防護起到重要的作用。
3 臨海環境腐蝕防護
海洋環境管線微生物腐蝕涉及材料與環境,因此,抑制和防止材料的微生物腐蝕就應該從這兩個方面來著手。目前,海底管道一般采用涂層保護、陰極保護、或是使用防腐鋼材和緩蝕劑殺菌劑的方式來進行微生物腐蝕的防護。
3.1 涂層保護
涂層保護是通過涂層的隔離作用來防止周圍環境中的腐蝕介質進入,抑制電化學、化學反應,以達到防腐的目的。良好的表面和優質的涂層會顯著提高深海裝備的涂層保護效果,有效抑制深海裝備的微生物腐蝕。涂層防護技術通常分內部防腐技術與外部防腐技術。
內部防腐技術主要針對管道內壁在油氣運輸過程中的內壁腐蝕的現象。為了防止管道內腐蝕,保障天然氣管道的安全,目前常見的防腐材料是環氧有機物質 (包括熔結環氧粉末與液體環氧涂料),其具有抗化學腐蝕性強、抗氧化、耐磨等多種優勢。
現階段聚乙烯材料是石油管道防腐常用的一種管道外涂層防腐材料,這種材料在具體應用過程中的優勢主要體現在涂層薄且附著力高、不透水、抗腐蝕、抗沖擊性能強、無毒等,同時,這種材料也具有較高的經濟性[50-52]。在一些管道規格變化較為頻繁的輸送管道施工中,常采用管道外壁纏繞聚乙烯材料的方式來對管道進行保護[53]。三層聚烯烴熱縮節點涂層已經成為國內外普遍采用的節點補口涂層結構。此外,針對冷管彎道進行防腐處理,可采用外層燒結環氧粉末制成涂層,以增強管道的抗腐蝕性能。在壓力較高的深海環境下,一般使用環氧瀝青、噴塑、聚烯等涂層用于深海裝備、管線防腐控制。
3.2 陰極保護
目前,大多數海洋設備都采用犧牲陽極的方法,其優點在于選材方便、安全可靠。這種方法的缺點是,海洋設備的防腐體系是處于一個動態的過程,陽極所能提供的保護電流及防腐涂層的狀態在不同時期不能確切地得知,也不能實現自動控制,通常為了保證防腐效果,其陽極的數量要足夠多。保護一旦停止,微生物附著引起的腐蝕將會對海下裝備造成非常嚴重的后果。此外,海洋環境中的微生物對犧牲陽極本身也會產生不利影響,一些研究表明海水中的SRB能夠直接或間接的促進犧牲陽極自身的溶解,從而減弱陰極保護效果。Liu等[13]研究了Al-Zn-In-Mg-Ti和Al-Zn-In-Sn兩種鋁基犧牲陽極在含SRB海泥中的腐蝕行為,實驗結果表明,大量的SRB附著在鋁基犧牲陽極表面,鋁陽極在含SRB海泥中的腐蝕速率明顯大于無菌海泥中的腐蝕速率且出現了更嚴重的點蝕。Guan等[54,55]研究了SRB誘導的Al-Zn-In-Cd犧牲陽極和5052鋁合金的腐蝕機理,附著在鋁合金表面的SRB能夠改變金屬材料和海水介質的界面特性,SRB參與電子傳遞過程并造成的陰極去極化效應加劇了鋁基犧牲陽極的腐蝕,且其腐蝕產物在SRB的作用下從Al(OH)3和Al2O3轉變為Al2S3和NaAlO2。因此,對于犧牲陽極的陰極保護而言,SRB能夠促進犧牲陽極的腐蝕從而減少犧牲陽極的使用壽命,在陰極保護系統中間接地影響了海洋管道的腐蝕。海洋輸油管道在服役期間還面臨一些高溫環境,在高溫條件下鋁合金犧牲陽極會出現電流效率降低,溶解不均勻等缺陷。在輸油管線高溫犧牲陽極中添加特定的稀土元素可以一定程度上解決犧牲陽極高溫電化學性能下降的缺點[56]。一些研究表明在高溫油氣環境下,Al-Zn-ln-Sn-Mg陽極具有良好的電化學活性,相比Al-Zn-In-Cd陽極而言具有更負的開路電位和驅動電壓[57]。外加電流陰極保護法是由外部直流電源直接向海洋設備上施加電流,使電子在金屬表面富集,并通過控制海洋設備的電位使海洋材料發生陰極極化,從而達到降低或抑制海洋設備腐蝕的目的。海洋管道外加的電流陰極保護系統可隨外界條件變化實現自動控制,使用周期更長。Guan等[58,59]研究了硫酸鹽還原菌和陰極保護共存情況下的高強鋼EQ70的腐蝕行為,結果表明,碳鋼在-0.85 V (vs CSE) 的保護電位下其腐蝕速率大于無陰極保護狀態,而在-1.05 V (vs CSE) 的狀態下腐蝕速率減小,且腐蝕產物由硫化物轉變為碳酸鹽,該研究對SRB存在條件下的陰極保護應用有很強的指導意義。
3.3 防腐鋼材與緩蝕劑殺菌劑
從材料角度出發,設計新型耐蝕材料可從本質上解決微生物的腐蝕問題。例如通過對材料表面進行合金化處理,對材料組織結構、力學性能實現優化,達到抗氫致開裂性能和耐微生物腐蝕性能的目的。通過在油氣集輸管道內壁加入緩蝕劑,可以有效降低內壁腐蝕問題的發生。加注緩蝕劑法是深海油氣管道的一種重要防腐措施。常用的緩蝕劑主要是含氮有機物。通常是在可能會發生腐蝕的位置或管道重要位置使用緩蝕劑,在相應區域形成穩定的蝕劑膜 (保護膜),將化學腐蝕物質與內壁進行有效隔離,達到防腐的目的,更好地對油氣集輸管道進行保護,提高輸油管道的運輸能力。當前海洋管道緩蝕劑的使用主要是為了防止海洋管道停運封存期間的內腐蝕,海管停運封存期間管道內部會充滿海水,微生物腐蝕會成為海管失效的重要原因。對于海洋管道腐蝕,目前已有各類綠色環保緩蝕劑的研究報道,鉬酸鹽類緩釋劑[60]、L-精氨酸-Zn+體系[61]、N-酰基肌氨酸衍生物[62]、聚天冬氨酸[63]等綠色環保緩蝕劑在海水環境中均展現出良好的緩釋效果,一些酶抑制劑還能夠抑制微生物的附著進而減少腐蝕的發生[64]。此外,殺菌劑也是控制微生物腐蝕的常用方法,Wang等[65]研究認為,吡啶硫酮鈉 (SPT) 不僅能夠在低濃度下有效抑制硫酸鹽還原菌的生長,而且SPT本身還具有明顯的緩釋效果,這種“一劑雙功能”的效果顯示出SPT優良的應用前景。
4 總結與展望
隨著海洋經濟的發展,微生物腐蝕導致的管線失效案例的增加,微生物對臨海環境管道造成的腐蝕問題日益受到重視。因此對臨海管道微生物腐蝕破壞行為、微生物腐蝕損傷機理及防護方法的研究將有利于全面了解海水管線的腐蝕情況,及時對管線腐蝕預警,避免發生更為嚴重的危害。目前雖然在微生物腐蝕規律方面做了大量研究,然而,實際海洋環境復雜多變,種類繁多的微生物在海水中共同存在,它們或彼此拮抗或彼此促進,相比于單獨存在的情況,微生物的代謝增殖特點必然會發生改變,海底長距離油氣集輸管線微生物腐蝕控制仍然是國家能源安全、國民經濟和社會發展的一個巨大挑戰。目前大多數的研究還集中在實驗室的研究,實驗室模擬實驗周期短,而微生物腐蝕是個長期的過程,因此要獲得對工業生產具有指導性意義的數據,需要大量實驗不斷積累經驗。此外,微生物的分布特征、腐蝕行為等因區域環境的差異而不同,導致微生物腐蝕規律復雜、腐蝕主要因素的分析與揭示仍是一大挑戰。針對這方面問題,需要對海水區 (飛濺區、潮差區)、海泥區 (潮差帶干濕交替區) 臨海管道的運行環境,采用現場分離的微生物,采用模擬與加速試驗方法,研究浪濺區、潮間帶區、海泥區管線材料微生物腐蝕損傷機理,研究重要影響因素對臨海管道微生物腐蝕的影響規律,建立微生物腐蝕模型及典型微生物腐蝕評價方法。
參考文獻
1 Ma S D, Li W H, Sun H Y, et al. The biological control of ocean corrosion [J]. Total Corros. Control, 2006, 20(3): 5
1 馬士德, 李偉華, 孫虎元等. 海洋腐蝕的生物控制 [J]. 全面腐蝕控制, 2006, 20(3): 5
2 Edyvean R G J. Biodeterioration problems of North Sea oil and gas production—A review [J]. Int. Biodeterior., 1987, 23: 199
3 Azis P K A, Al-Tisan I, Sasikumar N. Biofouling potential and environmental factors of seawater at a desalination plant intake [J]. Desalination, 2001, 135: 69
4 Yan T, Yan W, Dong Y, et al. Marine fouling of offshore installations in the northern Beibu Gulf of China [J]. Int. Biodeterior. Biodegrad., 2006, 58: 99
5 Heitz E, Flemming H C, Sand W. Microbially Influenced Corrosion of Materials [M]. Berlin: Springer-Verlag, 1996
6 Ma C, Chen C G, Jiang X B, et al. Distribution characteristics of marine bacteria in the China seas [J]. Med. J. Chin. PLA, 2012, 37: 909
6 馬聰, 陳昌國, 蔣學兵等. 中國海域海洋細菌分布特征分析 [J]. 解放軍醫學雜志, 2012, 37: 909
7 Liu H W, Xu D K, Dao A Q, et al. Study of corrosion behavior and mechanism of carbon steel in the presence of Chlorella vulgaris [J]. Corros. Sci., 2015, 101: 84
8 Duan L N, Liu Q Y, Jia S J, et al. Microstructure characteristics and strength-toughness of X100 pipeline steel [J]. Chin. J. Mater. Res., 2012, 26: 443
8 段琳娜, 劉清友, 賈書君等. X100級管線鋼的組織和強韌性 [J]. 材料研究學報, 2012, 26: 443
9 Dou W W, Jia R, Jin P, et al. Investigation of the mechanism and characteristics of copper corrosion by sulfate reducing bacteria [J]. Corros. Sci., 2018, 144: 237
10 Dong S, Bai X Q, Yuan C Q. Analysis of induced corrosion by fouling organisms on offshore platform and its research progress [J]. Mater. Prot., 2018, 51: 116
10 董碩, 白秀琴, 袁成清. 海洋平臺污損生物誘導腐蝕分析及其研究進展 [J]. 材料保護, 2018, 51: 116
11 Zheng J Y. Influence of marine biofouling on corrosion behaviour [J]. J. Chin. Soc. Corros. Prot., 2010, 30: 171
11 鄭紀勇. 海洋生物污損與材料腐蝕 [J]. 中國腐蝕與防護學報, 2010, 30: 171
12 Li H B, Zhou E Z, Ren Y B, et al. Investigation of microbiologically influenced corrosion of high nitrogen nickel-free stainless steel by Pseudomonas aeruginosa [J]. Corros. Sci., 2016, 111: 811
13 Liu F L, Zhang J, Sun C X, et al. The corrosion of two aluminium sacrificial anode alloys in SRB-containing sea mud [J]. Corros. Sci., 2014, 83: 375
14 Liu F L. Effect of sulphate reducing bacteria on corrosion of Zn, Al sacrificial anode materials in marine sediment [D]. Chongqing: Chongqing University, 2010
14 劉奉令. 海泥中硫酸鹽還原菌對鋅、鋁犧牲陽極材料的腐蝕影響研究 [D]. 重慶: 重慶大學, 2010
15 Liu F L, Zhang S T, Zhang J, et al. Effects of SRB on corrosion of pure zinc anode in marine sediment [J]. Chin. J. Mater. Res., 2010, 24: 411
15 劉奉令, 張勝濤, 張杰等. 海泥中SRB對純鋅陽極腐蝕行為的影響 [J]. 材料研究學報, 2010, 24: 411
16 Li Y C, Xu D K, Chen C F, et al. Anaerobic microbiologically influenced corrosion mechanisms interpreted using bioenergetics and bioelectrochemistry: A review [J]. J. Mater. Sci. Technol., 2018, 34: 1713
17 Videla H A, Swords C, Edyvean R G J. Features of SRB-induced corrosion of carbon steel in marine environments [A].
17 Dean S, Delgadillo G, Bushman J. Marine Corrosion in Tropical Environments [M]. West Conshohocken, PA: ASTM International, 2000: 270
18 Craig B D, McNeil M B, Little B J. Discussion of “mackinawite formation during microbial corrosion” [J]. Corrosion, 1991, 47: 329
19 Liu H F, Liu T, Zheng B J, et al. Influence of EPS's activity on 13Cr stainless steel's pitting sensitivity [J]. J. Huazhong Univ. Sci. Techno. (Nat. Sci. Ed.), 2009, 37: 122
19 劉宏芳, 劉濤, 鄭碧娟等. EPS活性對13Cr鋼鈍化膜點蝕敏感性的影響 [J]. 華中科技大學學報 (自然科學版), 2009, 37: 122
20 Stadler R, Wei L, Fürbeth W, et al. Influence of bacterial exopolymers on cell adhesion of Desulfovibrio vulgaris on high alloyed steel: Corrosion inhibition by extracellular polymeric substances (EPS) [J]. Mater. Corros., 2010, 61: 1008
21 Ghafari M D, Bahrami A, Rasooli I, et al. Bacterial exopolymeric inhibition of carbon steel corrosion [J]. Int. Biodeterior. Biodegrad., 2013, 80: 29
22 Chan K Y, Xu L C, Fang H P. Anaerobic electrochemical corrosion of mild steel in the presence of extracellular polymeric substances produced by a culture enriched in sulfate-reducing bacteria [J]. Environ. Sci. Technol., 2002, 36: 1720
23 Jin J T, Guan Y T. The mutual co-regulation of extracellular polymeric substances and iron ions in biocorrosion of cast iron pipes [J]. Bioresour. Technol., 2014, 169: 387
24 Xu D K, Li Y C, Gu T Y. Mechanistic modeling of biocorrosion caused by biofilms of sulfate reducing bacteria and acid producing bacteria [J]. Bioelectrochemistry, 2016, 110: 52
25 Xu D K, Li Y C, Song F M, et al. Laboratory investigation of microbiologically influenced corrosion of C1018 carbon steel by nitrate reducing bacterium Bacillus licheniformis [J]. Corros. Sci., 2013, 77: 385
26 Gu T Y, Jia R, Unsal T, et al. Toward a better understanding of microbiologically influenced corrosion caused by sulfate reducing bacteria [J]. J. Mater. Sci. Technol., 2019, 35: 631
27 Xu D K, Gu T Y. Carbon source starvation triggered more aggressive corrosion against carbon steel by the Desulfovibrio vulgaris biofilm [J]. Int. Biodeterior. Biodegrad., 2014, 91: 74
28 Zhang P Y, Xu D K, Li Y C, et al. Electron mediators accelerate the microbiologically influenced corrosion of 304 stainless steel by the Desulfovibrio vulgaris biofilm [J]. Bioelectrochemistry, 2015, 101: 14
29 Gu T Y. New Understandings of biocorrosion mechanisms and their classifications [J]. J. Microb. Biochem. Technol., 2012, 4: 1
30 Jia R, Tan J L, Jin P, et al. Effects of biogenic H2S on the microbiologically influenced corrosion of C1018 carbon steel by sulfate reducing Desulfovibrio vulgaris biofilm [J]. Corros. Sci., 2018, 130: 1
31 Chen Y J, Howdyshell R, Howdyshell S, et al. Characterizing pitting corrosion caused by a long-term starving sulfate-reducing bacterium surviving on carbon steel and effects of surface roughness [J]. Corrosion, 2014, 70: 767
32 Liu H W, Xu D K, Wu Y N, et al. Research progress in corrosion of steels induced by sulfate reducing bacteria [J]. Corros. Sci. Prot. Technol., 2015, 27: 409
32 劉宏偉, 徐大可, 吳亞楠等. 微生物生物膜下的鋼鐵材料腐蝕研究進展 [J]. 腐蝕科學與防護技術, 2015, 27: 409
33 Torres C I, Marcus A K, Lee H S, et al. A kinetic perspective on extracellular electron transfer by anode-respiring bacteria [J]. FEMS Microbiol. Rev., 2010, 34: 3
34 Reguera G, McCarthy K D, Mehta T, et al. Extracellular electron transfer via microbial nanowires [J]. Nature, 2005, 435: 1098
35 Liu H W, Gu T Y, Asif M, et al. The corrosion behavior and mechanism of carbon steel induced by extracellular polymeric substances of iron-oxidizing bacteria [J]. Corros. Sci., 2017, 114: 102
36 Wang H, Ju L K, Castaneda H, et al. Corrosion of carbon steel C1010 in the presence of iron oxidizing bacteria Acidithiobacillus ferrooxidans [J]. Corros. Sci., 2014, 89: 250
37 Liu H W, Liu H F. Research progress of corrosion of steels induced by iron oxidizing bacteria [J]. J. Chin. Soc. Corros. Prot., 2017, 37: 195
37 劉宏偉, 劉宏芳. 鐵氧化菌引起的鋼鐵材料腐蝕研究進展 [J]. 中國腐蝕與防護學報, 2017, 37: 195
38 Liu H W, Gu T Y, Zhang G A, et al. The effect of magneticfield on biomineralization and corrosion behavior of carbon steel induced by iron-oxidizing bacteria [J]. Corros. Sci., 2016, 102: 93
39 Liu H W, Fu C Y, Gu T Y, et al. Corrosion behavior of carbon steel in the presence of sulfate reducing bacteria and iron oxidizing bacteria cultured in oilfield produced water [J]. Corros. Sci., 2015, 100: 484
40 Hehemann R F. Stress corrosion cracking of stainless steels [J]. Metall. Trans., 1985, 16A: 1909
41 Xiong F P, Wang J L, Ahmed A F, et al. Research progress of sulfate-reducing bacteria induced SCC [J]. Corros. Sci. Prot. Technol., 2018, 30: 213
41 熊福平, 王軍磊, Ahmed A F等. 硫酸鹽還原菌誘導應力腐蝕開裂研究進展 [J]. 腐蝕科學與防護技術, 2018, 30: 213
42 Liu Q, Li Z, Liu Z Y, et al. Effects of H2S/HS- on stress corrosion cracking behavior of X100 pipeline steel under simulated sulfate-reducing bacteria metabolite conditions [J]. J. Mater. Eng. Perform., 2017, 26: 2763
43 Li X, Xie F, Wang D, et al. Effect of residual and external stress on corrosion behaviour of X80 pipeline steel in sulphate-reducing bacteria environment [J]. Eng. Fail. Anal., 2018, 91: 275
44 Zhou C S, Zheng S Q, Chen C F, et al. The effect of the partial pressure of H2S on the permeation of hydrogen in low carbon pipeline steel [J]. Corros. Sci., 2013, 67: 184
45 Xie F, Li X, Wang D, et al. Synergistic effect of sulphate-reducing bacteria and external tensile stress on the corrosion behaviour of X80 pipeline steel in neutral soil environment [J]. Eng. Fail. Anal., 2018, 91: 382
46 Biezma M V. The role of hydrogen in microbiologically influenced corrosion and stress corrosion cracking [J]. Int. J. Hydrogen Energy, 2001, 26: 515
47 Wu T Q, Xu J, Sun C, et al. Microbiological corrosion of pipeline steel under yield stress in soil environment [J]. Corros. Sci., 2014, 88: 291
48 Kennell G F, Evitts R W, Heppner K L. A critical crevice solution and IR drop crevice corrosion model [J]. Corros. Sci., 2008, 50: 1716
49 Laycock N J, Stewart J, Newman R C. The initiation of crevice corrosion in stainless steels [J]. Corros. Sci., 1997, 39: 1791
50 He T, Jańczewski D, Jana S, et al. Efficient and robust coatings using poly (2-methyl-2-oxazoline) and its copolymers for marine and bacterial fouling prevention [J]. J. Polym. Sci., 2016, 54A: 275
51 Banerjee I, Pangule R C, Kane R S. Antifouling coatings: recent developments in the design of surfaces that prevent fouling by proteins, bacteria, and marine organisms [J]. Adv. Mater., 2011, 23: 690
52 Liu J H, Qian S Q. Marine bioadhesion and defenses [J]. Corros. Prot., 2010, 31: 78
52 劉繼華, 錢士強. 海洋生物附著及其防護技術 [J]. 腐蝕與防護, 2010, 31: 78
53 Selim M S, Shenashen M A, El-Safty S A, et al. Recent progress in marine foul-release polymeric nanocomposite coatings [J]. Prog. Mater. Sci., 2017, 87: 1
54 Guan F, Zhai X F, Duan J Z, et al. Influence of sulfate-reducing bacteria on the corrosion behavior of 5052 aluminum alloy [J]. Surf. Coat. Technol., 2017, 316: 171
55 Guan F, Duan J, Zhai X, et al. Interaction between sulfate-reducing bacteria and aluminum alloys-Corrosion mechanisms of 5052 and Al-Zn-In-Cd aluminum alloys [J]. J. Mater. Sci. Technol., 2020, 36: 55
56 Liao H X, Qi G T, Yu K X. Research on sacrificing anode of high-temperature Al alloy containing rare earth and application [J]. Corros. Prot. Petrochem. Ind., 2004, 21(4): 19
56 廖海星, 齊公臺, 喻克雄. 含稀土高溫鋁合金犧牲陽極的研究與應用 [J]. 石油化工腐蝕與防護, 2004, 21(4): 19
57 Sun Y L, Wang N, Zhou Y, et al. Electrochemical performance evaluation of high temperature sacrificial anode under well environment [J]. Hot Work. Technol., 2017, 46(14): 99
57 孫雨來, 王楠, 周勇等. 油井環境中高溫犧牲陽極的電化學性能評價 [J]. 熱加工工藝, 2017, 46(14): 99
58 Guan F, Zhai X F, Duan J Z, et al. Influence of sulfate-reducing bacteria on the corrosion behavior of high strength steel EQ70 under cathodic polarization [J]. PLoS One, 2016, 11: e0162315
59 Guan F, Zhai X F, Duan J Z, et al. Progress on influence of cathodic polarization on sulfate-reducing bacteria induced corrosion [J]. J. Chin. Soc. Corros. Prot., 2018, 38: 1
59 管方, 翟曉凡, 段繼周等. 陰極極化對硫酸鹽還原菌腐蝕影響的研究進展 [J]. 中國腐蝕與防護學報, 2018, 38: 1
60 Wei Y L, Tian Y Q, Wang Y H, et al. Study on corrosion inhibition of compound corrosion inhibitor of molybdate in seawater [J]. Adv. Mater. Res., 2012, 581/582: 755
61 Gowri S, Sathiyabama J, Rajendran S. Corrosion inhibition effect of carbon steel in sea water by L-arginine-Zn2+ system [J]. Int. J. Chem. Eng., 2014, 2014: 607209
62 Kaskah S E, Pfeiffer M, Klock H, et al. Surface protection of low carbon steel with N-acyl sarcosine derivatives as green corrosion inhibitors [J]. Surf. Interfaces, 2017, 9: 70
63 Ma X M, Qian B, Zhang J, et al. The inhibition effect of polyaspartic acid and its mixed inhibitor on mild steel corrosion in seawater wet/dry cyclic conditions [J]. Int. J. Electrochem. Sci., 2016, 11: 3024
64 Liu F, Zhang L, Yan X, et al. Effect of diesel on corrosion inhibitors and application of bio-enzyme corrosion inhibitors in the laboratory cooling water system [J]. Corros. Sci., 2015, 93: 293
65 Wang J L, Hou B S, Xiang J, et al. The performance and mechanism of bifunctional biocide sodium pyrithione against sulfate reducing bacteria in X80 carbon steel corrosion [J]. Corros. Sci., 2019, 150: 296
免責聲明:本網站所轉載的文字、圖片與視頻資料版權歸原創作者所有,如果涉及侵權,請第一時間聯系本網刪除。
相關文章

官方微信
《中國腐蝕與防護網電子期刊》征訂啟事
- 投稿聯系:編輯部
- 電話:010-62316606-806
- 郵箱:fsfhzy666@163.com
- 中國腐蝕與防護網官方QQ群:140808414
點擊排行
PPT新聞

“海洋金屬”——鈦合金在艦船的
點擊數:7130

腐蝕與“海上絲綢之路”
點擊數:5741