
陰極極化對硫酸鹽還原菌腐蝕影響的研究進展
2021-07-22 16:31:34
作者:管方,翟曉凡,段繼周,侯保榮 來源:中國腐蝕與防護學報
分享至:


微生物腐蝕是微生物通過自身的生命活動直接或者通過其代謝產物間接的影響金屬材料腐蝕過程的現象。全世界每年因腐蝕造成的損失約占國民生產總值的3%~5%以上。硫酸鹽還原菌 (SRB)、鐵細菌、鐵氧化細菌、硫氧化細菌等微生物為重要元兇,一半以上 (50%~80%) 的地下管線腐蝕由微生物引起[,許多國家將15%~20%的工程費用用于防腐。2016年,全國腐蝕調查顯示,我國腐蝕成本占全部GDP的3.34%,總額超過21000億元人民幣。
微生物附著在工程材料表面形成生物膜,在材料和微生物膜界面處pH值、溶解氧、有機物和無機物等因素都與海洋本體環境完全不同。微生物活性可通過以下方式控制材料表面電化學腐蝕過程:(1) 微生物代謝過程影響材料的陰、陽極反應過程;(2) 微生物的代謝產物影響材料的陰陽極反應;(3)微生物通過在材料表面生成生物膜改變材料表面腐蝕環境;(4) 微生物直接參與金屬的腐蝕過程。陰極保護技術作為最有效的腐蝕防護方法已經得到世界范圍的承認,它被廣泛地作為油氣管道、船舶、海洋鉆井平臺等鋼鐵構筑物的腐蝕防護技術。陰極極化電位的施加改變了已有金屬與微生物之間的平衡,引起金屬-溶液界面性質變化,同時也影響著微生物的活動和金屬的腐蝕行為。本文結合SRB的生理特征和金屬材料陰極保護的可靠性,從陰極極化電位和 SRB 相互作用關系方面系統闡述和總結了施加陰極極化電位對SRB腐蝕的影響。
1 SRB的生態和生理特征
SRB是指一類能夠把硫酸鹽、亞硫酸鹽、硫代硫酸鹽等硫化物以及 S 還原成 H2S 的細菌的統稱。SRB廣泛存在于土壤、海水、河水、地下管道、油氣井等處。由于各地土壤類型、氣候狀況、地理位置等因素的差異,土壤中的SRB含量差別較大。我國庫爾勒、格爾木、拉薩、成都、大慶、沈陽、大港、鷹潭8個土壤試驗站的表層土壤的SRB含量小于或等于10~1800 cell·g-1不等。浙江省象山港海域表層海水和上覆水的 SRB 含量的年度均值分別為 173 和1000 cell·mL-1,沉積物中為 1440 cell·g-1;膠州灣潮間帶和沿岸區、克萊德海表層沉積物中的SRB含量則分別高達4.1×107和2.7×107 cell·g-1。大部分陸生SRB是中溫菌,其最適宜生長溫度為30~40 ℃;海洋中SRB的最適宜生長溫度稍低一些。SRB并不是嚴格的厭氧菌,分離自長慶油田采油回收污水水樣的 SRB 可耐受 4.5 mg·L- 1濃度的溶解氧。適合SRB生長的pH值范圍較廣,在5.5~9.0的酸堿度范圍內均能生長繁殖,其中7.5是比較合適的酸堿度條件。
SRB是最早的通過電子傳遞耦合磷化作用而非光合作用來產生三磷酸腺苷 (ATP) 的厭氧細菌,在此過程中SRB利用SO42-作為最終電子受體,從H或者有機酸的活化來獲得電子,同時產生腐蝕性S2-和有機酸等代謝產物。SRB 還原 SO42-的過程由一系列酶促反應組成。在這些酶促反應中,S得到8個電子,產生多個中間產物。硫酸鹽在硫酸腺苷轉移酶的作用下,以消耗ATP為代價激活硫酸鹽,使之生成腺嘌呤磷酰硫酸鹽 (APS)。反應生成的焦磷酸可被繼續水解,并由此促進整個反應的進行。相應的反應式如下:

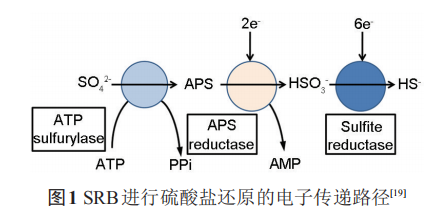
2 含SRB環境中金屬材料陰極保護的可靠性
要達到基本的保護需要-0.85 VCSE的保護電位(英國陰極保護標準)。Horvarth 和Novak于1964年提出,若水溶液中存在 SRB,則其陰極保護電位需要降低到-0.95 VCSE (即-1.022 VSCE) 的陰極保護標準。1981年,Fisher進行了實驗證實。通常隨著陰極極化電位的增加,腐蝕速率降低。要達到零腐蝕,理想的狀態是極化陽極達到它的可逆電位。在可逆電位狀態下,陽極溶解速率等于陽極沉積速率,沒有金屬的損失。但從實踐和經濟的角度看,腐蝕速率降低到10 mA/m2 (即11.6 μm/a) 時就可以被認為是得到了充分保護。保護不足和過保護都是不希望得到的狀態。過保護狀態下,可能引起快速的氫析出,從而導致鋼結構的氫脆。
陰極極化對金屬的SRB腐蝕行為的影響一直是研究的重點。在SRB存在條件下,最佳陰極保護電位負移。在鐵細菌中,負于-0.85 VCSE的陰極極化電位能明顯減少鐵細菌的數目,并達到較好的陰極保護效果。但是,在含SRB的介質中,陰極極化電位需達到-1.35 VCSE才能獲得相同的陰極保護效果。在含SRB的海泥中,對應-0.95,-1.03和-1.16 VCSE的陰極極化電流密度分別是5,11和82 mA/m2,相應的剩余腐蝕速率分別為12.4,8.9和5.5 mA/m2。在含SRB介質中,硫化鐵產物的不穩定保護作用是導致最佳陰極保護電位降低的重要原因。在Starosvetsky等研究中,在-0.1 VSCE的電位下,陰極表面的鐵硫化物沉積即可被還原。作者認為,若Fe的腐蝕發生在低于該電位的條件下,那么鐵硫化物的還原可能提供了一種新型的陰極去極化機理。陰極極化條件下,金屬表面的電位遠遠低于該電位,經由鐵硫化物還原導致的陰極去極化作用可能在金屬的腐蝕中發揮了重要作用。
3 陰極極化技術對SRB腐蝕的影響
3.1 陰極極化對金屬材料氫脆和力學性能的影響
在苛刻的海洋腐蝕環境中,對金屬構筑物進行陰極保護的腐蝕控制時,在金屬和海水界面會發生氧化還原反應,陰極反應為吸氧反應還是析氫反應取決于溶液中溶解氧含量、電位、pH值等。目前,設計時采用不正于-0.85 VCSE的陰極保護判據;當金屬所處環境中含有SRB時,則需要-0.95 VCSE甚至更負的電位。在更負的陰極極化過程中,金屬表面會逐漸形成鈣鎂沉積層,從而使材料的氫擴散系數降低,然而較高的陰極保護電位可能會對金屬產生過保護:
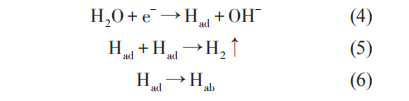
式中,Had和 Hab分別代表吸附在金屬表面和擴散到金屬內部中的H。
陰極反應所產生的H部分以H2的形式逸出,部分則吸附在材料表面,逐漸滲透到金屬的內部,從而引起氫脆,導致材料性能驟減而失效。常娥等對921A鋼的氫脆敏感性研究表明,在-0.96 VSCE極化電位下,陰極反應產生的Had進入材料內部,材料強度增加,韌性降低,耐氫脆應力腐蝕性能降低。當負于-1.01 VSCE以后,析氫反應加劇,出現了脆性斷裂。Yu 等研究認為,當金屬表面有 SRB 出現時,SRB 生物膜可以從極化電極中獲得電子傳遞給 H+催化產氫。此種情況下,吸附的H含量通常顯著增加,SRB存在時氫滲透電流密度是無SRB存在時氫滲透電流密度的3~4倍。
同時,一系列研究表明,SRB代謝產生的硫化物能增加金屬對H的吸附和滲透作用,進而導致氫脆的產生,但該促進作用因金屬的不同而存在一定的差異。相較于無菌和無H2S 的條件,SRB存在時低合金鋼的氫滲透略微增加;但對高強鋼和低合金鋼,SRB能顯著增加金屬表面生物膜中的硫化物濃度,從而促進金屬對H的吸收。Lunarska等的研究進一步證實,在含 SRB 的環境中,陰極保護條件下金屬表面弱保護性沉積物的形成、S2-的產生可能促進了鋼鐵材料的氫吸收。陰極極化條件下,SRB 對不同鋼材腐蝕行為的影響不同,其促進作用與 S2-的形成和硫化物沉積的形成有關。相同極化電位下,相較于波來鐵,糙斑鐵能吸收更多的H,且劣化也更為嚴重。總體來說,當海水中存在SRB 時,陰極保護應避免鋼材構筑物的某些部分處于-1.1~-1.2 VNCE的極化電位下。
國內一些學者研究了不同陰極極化電位對咸水環境 (3.5%(質量分數) NaCl溶液) 中Q235鋼氫脆敏感性的影響,并對斷口形貌進行了分析。結果表明,隨著陰極保護電位的負移,Q235鋼的力學性能 (最大抗拉強度和屈服強度) 并沒有呈現規律性變化,但斷面收縮率減小,材料發生氫脆的可能性加大。在-1.1 VSCE的陰極保護電位下,斷口出現了準解理斷裂特征形貌。
3.2 陰極極化對金屬構筑物周圍環境的影響
海洋環境中的金屬設施通常處于陰極保護狀態下。所施加陰極電流的大小與金屬表面所形成膜的厚度、結構、組成等有關。在陰極極化狀態下形成的腐蝕產物組成、鈣質沉積層和生物膜等性質與陰極電位大小有關。陰極極化使被保護金屬的電位整體負移,避免不同相間發生原電池腐蝕,從而導致在金屬表面形成鈣鎂沉積層和微生物膜。沉積層出現后可以抑制海水中的氧擴散到鋼表面,并提高陰極保護效率。然而,生物膜的附著和鈣質層的形成也使得極化電阻增大,要達到相同的電位所需極化電流增大。
當對被保護金屬施加陰極電位時,在陰極極化的初始階段,O2被還原,生成OH-:
在更負的陰極極化電位下,H2O 被還原生成OH-和H2。
以上反應表明,隨著陰極極化電位的負移,金屬材料周圍環境中的pH值升高,環境逐漸變為堿性,并導致鈣質層的沉積。
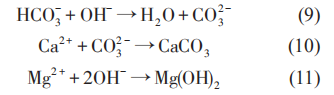
陰極極化條件下生物膜的結構特點和腐蝕產物 (無機沉積) 發生了變化。Sun等研究了陰極極化條件下,土壤中 SRB 對 Q235 鋼腐蝕行為的影響,認為當陰極極化電位從開路電位 (約-0.82 VCSE)負移至-0.85 VCSE時,其 pH 值約升高 1;但隨著陰極極化電位繼續負移時,SRB 數量減少,但由于 CO32-和Ca2+的緩沖作用,pH值變化不大。同時,Ca2+濃度減小和pH值升高可以顯著抑制細菌的附著。使用X射線能譜 (EDX) 對陰極保護條件下Q235鋼表面的腐蝕產物組成進行分析表明,在不含SRB的土壤中,Q235 鋼表面的腐蝕產物中沒有 FeS;而在含SRB 的土壤中,Q235 鋼表面則覆蓋有一層黑色的FeS。在自腐蝕電位下,腐蝕產物中不含C;而在極化電位下,由于陰極極化電流降低了HCO3-與Ca2+反應生成CaCO3的活化能,無論有無SRB存在,腐蝕產物中都含C。
當鋼鐵材料浸泡在細菌介質中時,表面可能形成各種類型的鐵硫化物。研究表明,腐蝕產物Fe1+xS開始時能為金屬提供保護,但是當該種產物層變厚時,有可能發生開裂,導致腐蝕速率增加。同時亦有研究表明,FeS 在細菌的胞外電子傳遞中起著重要的作用,HS-和可溶性聚硫化物在將電子從希瓦氏菌 (Shewanella oneidensis) MR-1胞外傳遞到與細菌接觸的FeS中起著重要作用。在含有SRB介質中,陰極極化電位的增加,使鐵的腐蝕產物由硫酸鹽向碳酸鹽發生轉變,并且隨著陰極極化電位的負移,轉化的碳酸鹽含量增加。
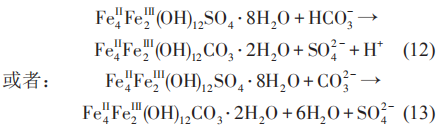
3.3 陰極保護對SRB的影響
在天然海水中,陰極極化電位的施加可有效抑制生物膜的附著,陰極氧還原是陰極極化抑菌的主要機制。然而對于無氧環境中,陰極極化作用未能有效抑制厭氧微生物的附著。陰極極化條件下,不銹鋼表面附著的SRB的數量未減少,該特點與好氧細菌生物膜不同。-0.95 VCSE的陰極極化電位下,微生物的活動未受影響,種群生長 (電解液環境中,非試片表面) 反而略微增強。雖然已有很多研究表明陰極保護下的生物膜和鈣質層之間有密切聯系,但是由于實驗室環境和實際海洋環境存在差異,海水中有機物的差異造成了電流密度、形成的鈣質層和生物膜的不同,要探究生物膜和陰極極化的關系并不容易。
陰極極化對SRB代謝影響可以分為以下幾種:
3.3.1 陰極極化抑制 SRB 代謝 陰極保護條件下,在極化電極表面產生自由H的速率高于SRB去極化過程中的速率。有活性SRB存在時,外加保護電位會降低0.1 V,即降到-0.95 VCSE,才可以產生保護作用。甚至有研究結果表明,由于陰極極化導致金屬腐蝕電位改變,從而在無菌環境中的陰極極化電位在SRB環境中變為陽極極化,即在SRB介質中金屬的開路電位比無菌環境中最佳陰極保護電位更負。
當金屬構筑物浸泡在海水中,微生物附著在金屬表面,吸附作用在微生物附著的初期 (約當浸泡在海水中之后的前 100 min 之內) 起主導作用。陰極電位施加之后,附著的細菌數目明顯減少,特別是細菌附著初期,這說明陰極極化電位的施加通過靜電作用影響了微生物的吸附過程,同時施加極化后在界面上產生H2O2以及多余的OH-等,抑制了微生物的附著。
陰極極化下 SRB 的代謝活性一直是研究者探討的熱點。Lunarska等研究在人工海水中陰極極化下SRB對金屬力學性能的影響時認為,在低弱陰極極化時,SRB 在電極表面產生 H2S,使得界面 pH值降低,抑制鈣質層的出現。但是在強陰極極化時,由于界面產生的強堿性區 (反應式 (6) 和 (7)) 使得界面pH值升高,抑制了SRB的生長。李國華等研究含SRB土壤中陰極極化對Q235鋼腐蝕的影響時認為,隨著陰極極化電位不斷負移,Q235鋼件周圍的土壤中SRB數量逐漸減少,但在-1.05 VCSE的陰極極化電位下,Q235鋼試件周圍土壤中SRB仍能夠存活;在相同的外加電位下,Q235鋼在有菌土壤中所需要的陰極極化電流密度較大。Dhar 等認為負于-1024 mV的陰極極化電位可以使微生物濃度降低1~2個數量級。施加脈沖陰極極化可同樣抑制微生物的活性,5~20µA·cm-2的陰極電流密度可以使微生物濃度降低2~3個數量級。將施加陰極極化的不銹鋼浸泡在 Vibrio alginolyticu 溶液中,陰極極化3 h能明顯減少微生物的附著,但是在浸泡時間超過9 h以后,在極化和未極化體系中微生物的差異消失。
3.3.2 陰極極化促進 SRB 代謝 SRB 代謝活性與陰極極化電位密切相關,隨著陰極極化電位的負移,SRB的數目呈現先增加后減小的趨勢,FeS的含量變化趨勢與 SRB 細菌數目一致。強陰極極化電位下SRB數目的減少與其界面pH值的升高密切相關,而弱陰極極化下SRB數目增加說明弱陰極極化電位促進了SRB的代謝活性。趙曉棟等研究了含有 SRB 的海泥中陰極極化電位對碳鋼腐蝕的影響,通過最大概率計數法結合不同陰極極化電位下的 Bode 圖得出,在低電位(-0.85 VCSE) 下細菌的生長活性與穩定性高于在較高陰極極化電位 (-0.95 和-1.05 VCSE) 下的,且高于無施加陰極極化電位下的細菌數目。對硫還原地桿菌 (Geobactersulfurreducens) 的研究表明,生物膜形態和結構對金屬表面的電流密度具有重要影響,離散的、稀疏的菌膜產生的電流密度高于致密的菌膜。因此不同陰極極化電位下,細菌生物膜形態和結構的改變及活性的變化都會對金屬的腐蝕產生一定的影響。SRB 作為一種電活性微生物,其與極化金屬間電子傳遞方式可分為直接電子傳遞 (DET) 和間接電子傳遞 (MET)。圖3顯示了SRB在與極化電極之間的DET和MET方式。
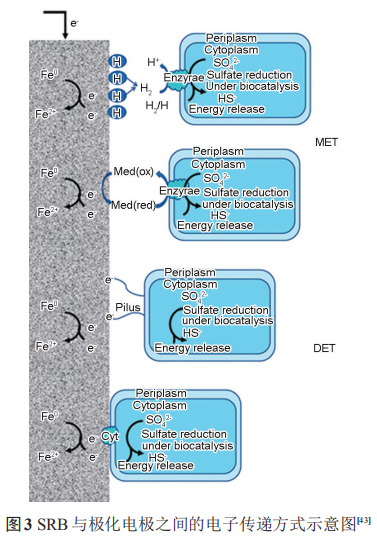
(1) 間接電子傳遞Fe 失去的電子須借助電子傳遞介質才能傳遞到SRB中。在微生物燃料電池中,電子傳遞介質的使用可以增加細菌的產電能力。電子傳遞介質從陰極極化的電極中獲得電子,通過電子傳遞通道,將電子傳遞到SRB中。H可以作為SRB與極化電極之間間接電子傳遞的介質。施加陰極極化后,H+向電極表面移動并在電極表面上還原成[H];而[H]在SRB中氫化酶的催化下還原SO42-并為SRB的代謝提供能量,從而促進SRB的生長代謝。同時,在此過程溶液的pH值升高。研究表明,Desulfitobacterium 可以通過在極化-0.7 VSHE電極上進行H的活化。產電微生物Geobacter sulfurreducens可以利用極化電極上H的氧化還原進行產電,其反應如下:
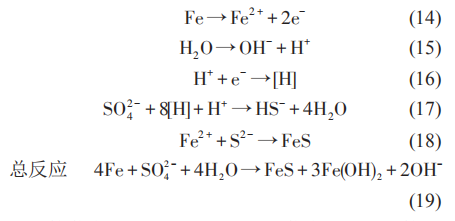
核黃素、奎寧胡敏酸、吩嗪和黃素腺嘌呤二核苷酸 (FAD) 等是常見的內生電子傳遞介質。SRB 可以利用維生素B12等與極化的電極進行間接的電子傳遞。在SRB介質中,鐵硫化物也可以作為SRB的電子傳遞通道。在這個過程中,硫酸鐵 (Fe2(SO4)3)可以作為 SRB 的電子傳遞受體,從而將鐵的腐蝕產物轉化為碳酸鐵 (Fe2(CO3)3)、藍鐵礦 (Fe3(PO4)2·(H2O)8)、硫化鐵 (Fe2S3) 等,同時促進 SRB 的代謝活性,如圖4所示。
(2) 直接電子傳遞直接電子傳遞為SRB利用細胞色素c等電子傳遞蛋白或者自身形成的納米線與電極直接進行電子傳遞。Sherar 等和Xu等的研究表明,在碳源不足的條件下,SRB會形成納米線從金屬基體中直接獲得電子。
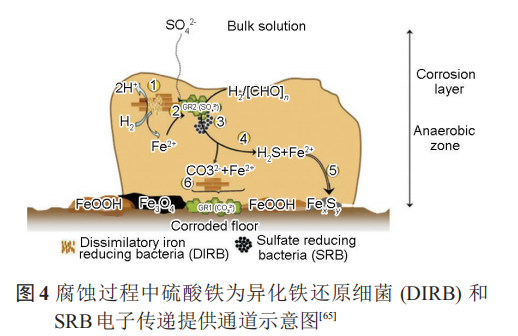
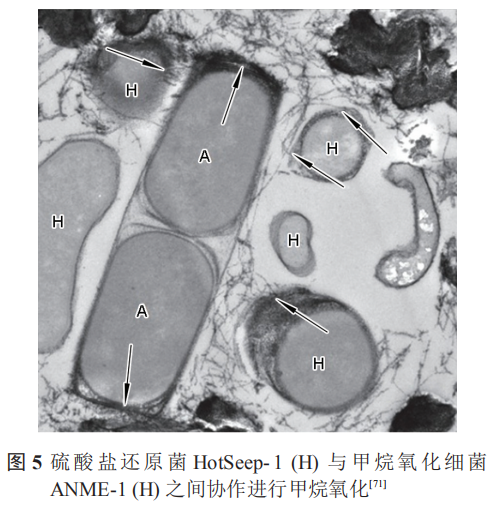
除此之外,在共生的細菌中,非產電細菌亦可為產電細菌提供電子通道。Wegener等證明甲烷氧化細菌 (ANME-1) 與 SRB (HotSeep-1) 之間可以通過形成納米線進行電子傳遞,圖5中箭頭明確指出了電子傳遞的納米線。
陰極極化電位影響SRB的電子傳遞方式,并影響著 SRB 的代謝活性。在合適的陰極極化電位下(如-0.4 VSCE或-0.7 VHSE等),SRB 可以從極化的電極中直接或間接獲得電子用于自身的代謝,即較低的陰極極化可以促進SRB的代謝,而強陰極極化電位則會抑制SRB的代謝和生長。極化作用所在電極表面產生的強堿性環境,并不利于細菌附著,這反而抑制了SRB的活性;此外,已有研究證明,金屬表面形成的FeS等腐蝕產物也為SRB從溶液鐵金屬中直接獲得電子提供了通道 (圖6)。
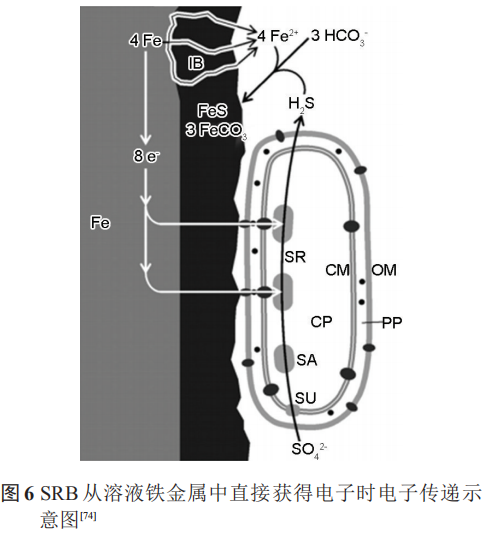
Nekoksa等研究認為,-1 VSCE的陰極極化使金屬表面pH值降低,金屬表面SRB數目增多。當電位低于-1 VSCE時,pH值逐漸呈現堿性,SRB數目降低。利用電化學測試和生物分析方法探究陰極極化下的微生物活動結果表明,-0.9 VAg/AgCl的陰極極化可以在短期內為海洋金屬設施提供保護;但是當極化終止后,厭氧SRB將通過消耗陰極極化過程中產生的H來促進自身的生長代謝,從而加速金屬的腐蝕。
4 問題與展望
近幾十年來,微生物腐蝕已經成為國內外腐蝕與防護研究的重點,人們在研究陰極極化與微生物間的相互作用方面也取得了很大的進步。但要全面深入地認識微生物腐蝕與腐蝕防護技術 (如陰極保護) 的相互作用,仍有諸多問題需要解決。如近些年的研究表明,生物膜內細菌群落組成的差異和細菌代謝活動產生的差異對微生物腐蝕具有重要影響,然而在評估生物膜對金屬腐蝕的影響時很少考慮到SRB的代謝特異性。2004年,Dinh等在只有Fe為電子受體的培養基中分離出脫硫弧菌屬 (Desulfobacterium) 的一個新種,相較于傳統的耗氫型菌株,該菌株還原硫酸鹽的速度更快,作者推測此類菌株能夠從金屬中直接獲得電子。Venzlaff等從實驗驗證了以上推測。Yu等從鐵銹中分離純化了Desulfovibrio caledoniensis,并證實其能夠從-0.74 VSHE的極化電極中獲得電子,即利用陰極作為電子供體,這種直接電子傳遞的通路在生物代謝和加速金屬腐蝕中有重要作用。其他的一些研究也表明,同一個屬的不同菌株對金屬腐蝕行為存在很大的不同。因此,在將來有關微生物腐蝕的研究中,加強物種多樣性的調查是非常必要的。對于那些能夠直接從金屬中獲得電子的SRB菌株,有關其腐蝕過程中電子傳遞機理方面的研究甚少。利用分子生物學、光譜電化學及微區腐蝕觀察等技術手段研究 SRB 菌株的直接電子傳遞途徑及其呼吸代謝機制,對未來微生物腐蝕研究探索有重要意義。
免責聲明:本網站所轉載的文字、圖片與視頻資料版權歸原創作者所有,如果涉及侵權,請第一時間聯系本網刪除。
相關文章

官方微信
《中國腐蝕與防護網電子期刊》征訂啟事
- 投稿聯系:編輯部
- 電話:010-62313558-806
- 郵箱:fsfhzy666@163.com
- 中國腐蝕與防護網官方QQ群:140808414
點擊排行
PPT新聞

“海洋金屬”——鈦合金在艦船的
點擊數:6534

腐蝕與“海上絲綢之路”
點擊數:5265